Elastic Matching and its Applications
Colin Windsor
Abstract
The smooth non-linear mapping between one multivariate function and another
which both preserves the local details at corresponding points in the map and
also minimises the complexity of the map is the basis of elastic matching. Its
roots in two—dimensional image mapping will be described, but many of its
methods are best appreciated in one dimension, which has several important
applications in its own right. The general procedures and methods for
enhancing success will be described together with applications to gas
chromatography, static and dynamic signature recognition, and face
recognition.
1. Introduction
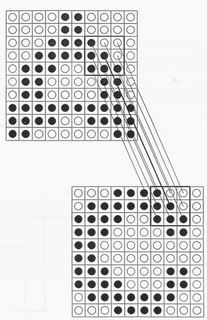
The biological basis of elastic matching was described by Beinenstock in terms
of the structure of the human visual cortex. Figure 1 after his paper describes
the type of connections which are thought to exist between different layers in
our visual cortex. When corresponding neurons on a template layer (above) and
test layer (below), indicated by the heavy line, have both a common firing mode
(black or white) and also a common set of nearest neighbour enviromnent of
firing pattems then a reinforcement mechanism is assumed to apply and the
connections between the two layers are reinforced. Figure 2 shows the type of
nearest neighbour neuronal circuit which Von der Marlsburg and Bienenstock
[2] describe. Two adjacent firing neurons (black) send their signals both to their
near neighbours, and to a second similar layer of neurons. A four-neuron loop
is therefore formed when both emitter and receptor neurons have the same
firing mode and so would tend through Hopfield’s Hebbian learning reinforce
the connections between them. The common nearest neighbour environment
may readily be generalised to include the more distant surroundings of the
central connection, and less precise matchings. In general a good match can be
specified by high correlations between the connected regions of the template
and test functions.
Figure 1. The type of links suggested to occur in aeyacent layers ofthe
human brain by Beinenstock[1]. Particular neurons in the two layer (as
joined by the full line) have several neighbours with similar pixels on each
layer, giving a series of reinforcing circuits.
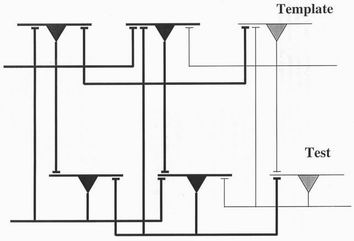
Figure 2. The neuronal picture of adjacent pixels (black or white) in the two
layers storing the prototype and test images. Links from the axon of each cell
connect both with adjacent layers and with near neighbours in the same
layer. A four-neuron circuit of neurons with a similar firing pattern can
therefore be established.
For example corresponding positions in a hand-written
signatures could include correlations between the spatial
positions, and also the pen timings, pen velocities, and tip pressures. This type
of correlation mapping is invariant to spatial displacements between the objects
and at least partly invariant to rotations and scale changes.
The second fundamental principle of a good mapping is that it should be
appropriately simple. In many practical cases nearly perfect local correlations
can be achieved between points in the two functions which are nevertheless
incorrectly matched. For example in signature recognition of the name "Bill
Full", the both words end with the same "...ll" sequence. As illustrated in figure
3, it is likely that there will be good local correlations from any one example in
the template to both examples in the test signature. The ambiguity can be
minimised by including more distant correlations, but it can more easily be
removed by requiring that the matching transformation between the two
signatures be a smooth one. The match to the correct position gives a smooth
function, while the match to the second requires a sudden gap.
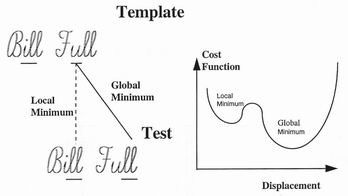
Figure 3. Two hand-written signatures being compared. Any correlations
existing between the marked "ll" sequence arebound to be echoed in the
false connection which therefore represents a local correlation minimum.
The name elastic matching derives from the technique of finding a good linear
match between two images by printing the test image on a transparent elastic
sheet, and stretching the sheet until the template and test images optimally
overlap. The technique is readily generalised to functions of higher or lower
dimensionality and to non-linear mappings of all kinds. The concept leads
naturally to a particular criterion for a possible mapping - that which leads to
the minimum elastic strain energy within the elastic sheet. According to
Hooke's law for a perfectly elastic medium, the force F when an elastic object
of length x is stretched by a length dx is F=Ydx,
and the elastic strain energy is
E=Ydx2/2. Consider a template function array xA(i)
compared to a test
function xB(j), connected by a link array L(i)
which connects the ith point on
the A array with the j=L(i)th point in the B array.
The total elastic energy is
then seen to be proportional to
E=Y Si
(xA(i)2 - xB(L(i))2 )/2. Thus the
minimum elastic energy corresponds to the minimum least squares differences
between the points on the template function and the corresponding matched `
points on the test function. Unfortunately such a simple algorithm is not
invariant to even translational displacement which will certainly stretch the
elastic sheet! Appropriate preprocessing to define objects with respect to their
centres of gravity, their principal axes, and their principal radii could restore to
the elastic sheet its objective of looking for a smooth set of matching
connections, but such an approach will never be optimal as it will always try to
remove any type of distortion between the functions. What is needed is a
function which is neutral to smooth elastic distortions but which penalises the
spatially sharp distortions produced by incorrect linkages.
2. Obtaining a good match - a one dimensional example
It is usually easy to define, as in the above section, the criteria by which a
satisfactory elastic match can be judged. It is even easier for the human eye to
judge a successful match, and note any inconsistencies. The problem is how to
achieve it.
An important one-dimensional example of the need for elastic matching occurs
in gas chromatography. A small sample of say peppermint to be assessed
against a standard sample is vaporised and the gas passed through a long
chromatograph whose output is a series of peaks describing the arrival time of
different molecular species. For the purpose of this discussion it is assumed that
the raw data is separated into a series of discrete peaks as similar height as a
function of the retention time through the chromatograph. Figure 4 shows such
a spectrum for a template sample A (upper) and a test sample B (lower).
The problems of connecting corresponding peaks in the two spectra are evident.
There is a considerable displacement between the main groupings of peaks
which the eye is quick to see in both patterns. However the displacement is not
constant or even linearly changing along the spectra. Some template spectrum
peaks have no corresponding partner in the test spectrum - we call these
"dangling" peaks. Some test spectrum peaks do not appear in the template -
"rogue" peaks. Some peaks in the template would like to connect to a single
peak in the test spectrum - "Multiple" peaks.
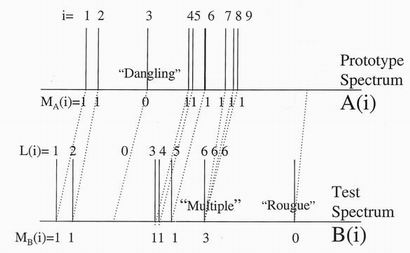
Figure 4. Two gas chromatography spectra being compared. Peaks i in the
prototype are linked in to peaks L(i) in the test. Various types of mismatch are
shown. The arrays NA(i) and NB(j)
will be used to define a cost energy.
It is instructive to consider how the eye functions when presented with such a
pair of spectra. First it searches for the most distinctive cluster of peaks —
distinctive because its central peak has correlations with those in its near
neighbourhood that are not common in the spectrum. The eye then searches in
the test spectrum for a similar cluster. If it finds one, then the eye mentally
draws the first link between the corresponding central peaks. The process can
then be repeated for the second most distinctive cluster of peaks. If the two
links are correct then an approximate mapping is established and if the mapping
is at all linear it will often be easy to add other links. If the mapping is more
non-linear then the process of searching for distinctive clusters of peaks will
have to be continued within the regions of the spectrum between the first links
identified.
The problems arise when some of the early links were incorrectly made. Lack
of any exact match, even with the correct links, can easily lead to an initially
incorrect assignment. This is where the eye brings in the elastic sheet. It is able S
to size up both spectra as a whole and favour those altemative links where the
local stretching of the sheet is minimised.
In a complex pair of spectra with many apparently similar local clusters, none .
perfectly matching, the skilled eye begins the process which our methods
attempts to simulate - that of trying out a wide range of possible connections,
weighing up the balance of local correlations and distortions, and slowly
settling down on those connections which show most promise.
3. The quantitative description of local correlation
The correlation and distortion concepts may be related by considering each as
an elastic energy. The correlation energy representing the elastic energy
required to map the environment of the central peak of one local cluster of
peaks in the template, when linked to that of another in the test spectrum. The
distortion energy is the contribution to the elastic energy of the total mapping
from the chosen linkage. .
Consider two sets of gas chromatography peak positions, a template spectrum
composed of NA peaks of intensity y A(i) at times
tA(i), and a test spectrum
with NB peaks yB(j) at times
tB(j). Suppose a current set of linkages between
the two spectra j=L(i). The correlation energy may then be written as illustrated
in figure 5 as
Ecorr = Si=1,NA
Sk=-ncorr,ncorr [ XA(i)
- XA(i+k) - XB(L(i)) + XB(L(i+k))]2
where the sum over i runs over all the peaks and the sum over k runs over the
ncorr peaks on either side of the peaks i in spectrum A or
L(i), the linked peak to
i in spectrum B. The correlation energy as so defined is proportional to the
elastic deformation energy needed to transform a map of the immediate
neighbourhood of a chosen peak in the prototype into that of its linked partner
in the test spectrum. Similar correlation terms can be defined for peak intensity
in gas chromatography, for variables such as the y position, the timing or
pressure of a pen stroke, or for the non—positional variables of a circular arc.
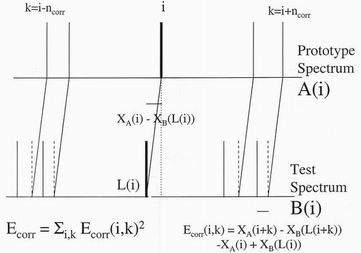
Figure 5. The correlation energy between two linked peaks. It is the sum over
neighbouring peaks i+k of the their displacements relative to the
displacement of the chosen peak i.
4. The quantitative description of local distortion
The distortion energy term needs to be related, not to the elastic energy of
distorting one spectrum into another, but to the extra elastic energy needed to
make the distortion over the best current measure of a smooth distortion
between them. For this purpose a distortion vector xD(i)
is defined which may
be evaluated for a given set of links from the average difference between linked
points over the immediate neighbourhood on either side of the ith peak.
The distortion energy term is linked to the extra elastic energy needed to distort
the points in the prototype spectrum into linked points in the test spectrum. The
first step in evaluating this energy is to calculated a "distortion vector"
averaged over the ndist lines on either side of the chosen line i
xD(i) = Sk=-ndist,ndist
xA(i+k) - xB(L(i+k)).
This array, together with a similar y array in two dimensional problems, can
then be used in the evaluation of a distortion energy
Edist = Si=1,NA
[ xA(i) - xB(L(i)) - xD(i)]2.
Such an energy term will then penalise the difference between the actual linked
ordinates on the two spectra and their average values.
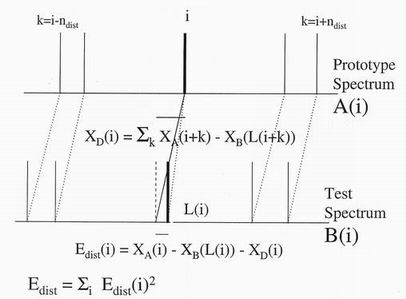
Figure 6. The distortion vector at peak i is the mean displacement over
neighbouring peaks i+k. The distortion energy between two linked peaks is
the sum of the squared differences between the actual displacements and the
distortion vectors.
5. Matching all the links
One of the tasks the eye is good at, is spotting, and making due allowance for,
the inevitable links that will never match. These include the "dangle" links
when peaks present in the prototype not matched by any in the test, unlinked
"rogue" peaks present only in the test and "multiple" links when two or more
peaks in the prototype connect to a single peak in the test. The matching
process must encourage a one—to-one mapping where best and allow for
dangle, rogue and multiple links when these are necessary. These objectives are
satisfied by defining two multiplicity arrays mA(i) and
mM(j) defining the
number of connected links to each point in the corresponding spectra and
defining a "bond" energy
Ebond = Si=1,NA
[ mA(i) - 1]2 -
Si=1,NB
[ mB(j) - 1]2.
This energy is zero for fully linked spectra with multiplicity 1 for all links but is
penalised by one unit for any dangle, rougue or double bond as was illustrated in figure 4.
The total energy function will now take the form
Etot = YcorrEcorr + YdistEdist +
YbondEbond
where the constants Y are adjustable parameters of the method. We wish to
reduce all terms to zero so the relative magnitudes of our multipliers may be
chosen at our discretion.
6. Allowed links in one, two and higher dimensional problems
Our example so far has been one dimensional, when all peaks are a function of
the single variable, retention time. There is then a fundamental restriction on
allowed links that they must preserve the time ordering in each spectrum
L(i) > L(j) if i>j.
A similar relationship may hold in two dimensional patterns dictated by a single
variable, such as time in dynamic signature verification. Corresponding points
along the two—dimensional signatures can strictly only be linked in such a way
that the time ordering is preserved. Experts to dynamic signature verification
. use test their systems with a simple cross. As illustrated in figure 7 it can be
drawn in 8 ways, and only one way is correct! Apparently identical signatures
can differ only in stroke order or direction. The strokes of a signature recorded
on paper without time information represent a truly two dimensional pattem. In —
this case there is no restriction on the connectivity of the links and all 8 crosses
are equivalent. However a pictorial image described by a static pixel array in
two dimensions and subject only to non—linear distortions will connectivity
between pixels which imposes a more complicated two-dimensional link
structure. A dynamic signature tablet including speed and pressure variables
will still have one dimensional connectivity.
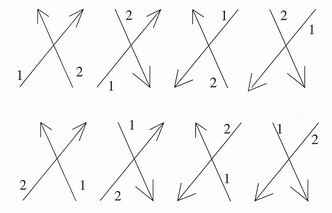
Figure 7. In a dynamic signature veryfication the timing and the order of
strokes imposes limitations on the connectivity of points in the traces. A
simple cross may ble drawn in 8 distinct ways.
7. Making a set of initial linkages
This is a critical step, since the problems of local minima mean that mistakes in
initialisation can be computationally expensive to correct. One method is to
mimic the eye by choosing in order the most distinctive features (eg local
pattems or largest peak intensity) in the prototype spectrum and making the
connections in this order. Alternatively only a fraction, say l/8 of the peaks,
should be linked. The optimum spacing between chosen peaks would be
comparable with the likely distortion. Such a selection of peaks is shown in
figure 8. A search for the best linkage can then be made by examining the
correlation energy only, as possible linkages are tried over the allowable range
given by the existing linkages on either side. The linkage with the lowest
correlation energy would be chosen. After match improving using all the
energy terms, as described in the next section, a new set of links could be added
between the first by doubling the number of chosen peaks, and repeating the
process until all the initial connections had been made. The process of altering
link positions is very much easier when only a fraction of linkages are defined,
than for a full set of linkages when existing links prohibit eachother’s
movements.
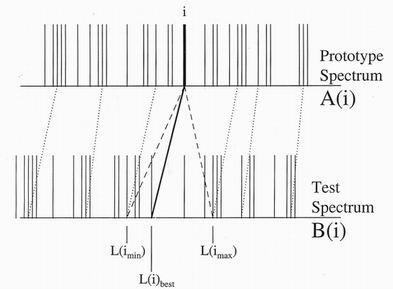
Figure 8. New links are best chosen for only a selection of possible peaks.
The optimum link to the chosen peak i can initially be found from that
within the allowed range with the lowest correlation energy. The selected
links can then be optimised before adding more linkages.
8. Improving the match: Monte Carlo method simulated annealing
Given a set of initial linkages, even a partial one, the problem is how to reduce
the total energy as far as possible. Assuming that changes in the linkages can be
made only by moving links one at a time, there may need to be many
intermediate unfavourable links before the best link is encountered. The total
energy surface thus contains local minima which must be overcome. Monte
Carlo simulated annealing is an old recipe for overcoming such problems. A
"thermal energy" kT is defined and possible link changes may be accepted even
when they increase the total energy. The recipe is that defined in 1952 by
Metropolis et al
(i) Changes are accepted if the energy change
DE due to the new linkage is less than zero
DE < 0
(ii) Changes are also accepted if the energy change is such that
exp(DE/kT) < rnd
where rnd is a random number between 0 and l.
The method eventually populates all possible states according to the Boltzmann
probability function exp(-DE/kT)
and so allows movements through an energy
maximum towards a global energy minimum provided that the thermal energy
is comparable with the barrier height. At such temperatures there is inevitably a
population of non—optimal linkages. The annealing process consists of slowly
reducing the thermal energy to zero so that all linkages slowly drop into their
lowest energy state. The magnitude and number of tries at high themal energies
must be related to the energy barriers required to be overcome.
Copyright 1997: Colin Windsor: Last updated 4/12/2010